Biostratigrafický výzkum jeskyně ve Skalce nad Čihovou

Biostratigraphic investigation of the cave in the hill Skalka nad Čihovou
Vojen Ložek
(Československý kras 38; str.55—69; Praha 1987)

V roce 1984 jsem provedl biostratigrafický výzkum výplně vchodu jeskyně v návrší Skalka nad Čihovou u Karlštejna. Na rozdíl od jiných dosud uveřejněných nalezišť obdobného vývoje a stáří (Ložek 1974, 1982a), která vesměs leží při dně údolních zářezů nebo na strmých skalnatých svazích, otvírá se ohledaná jeskyně při úpatí svahu oblého návrší na mírně zvlněné parovinné plošině, daleko od ostrých erozních tvarů. Jde tedy o polohu, z níž dosud nemáme přesnější doklady ani o vývoji fauny, ani o průběhu sedimentace a odnosu. Diverzita biocenóz i stanovišť je zde sice daleko nižší než ve zmíněných členitých okrscích, na druhé straně se však právě taková poloha blíží klimaxovým poměrům, což má značný význam, jak pro poznání vývoje nejmladšího kvartéru, tak z hlediska ochrany přírody, neboť jeskyně leží v rezervaci Karlštejn. Naše poznatky mohou přispět k řešení těchto otázek:
- Vývoj postglaciální fauny v parovinném prostředí o nízké diverzitě, zejména ve srovnání s poměry v údolních zářezech.
- Sledování akumulačních a odnosných pochodů i vývoje sedimentů v uvedeném prostředí.
- Sledování půdotvorných pochodů, především ve vztahu k sedimentaci, odnosu a krasovění.
- Rekonstrukce celkových přírodních poměrů v tomto úseku rezervace a využití získaných poznatků při ochranářské péči o plochy obdobného rázu.
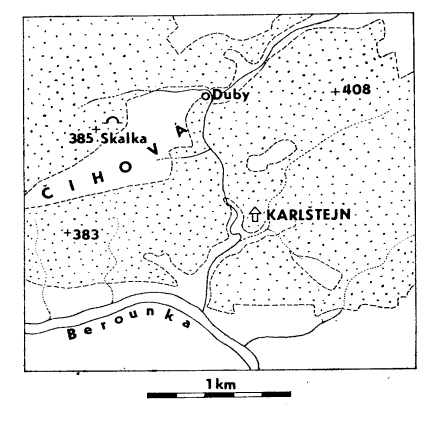
Poloha jeskyně ve Skalce nad Čihovou. – Location of the Skalka Cave.
Poloha jeskyně a popis profilu v jejím vchodu
Jeskyně se otvírá při úpatí s. svahu návrší Skalka 1,25 km na SZ od hradu Karlštejna. Základní údaje o její poloze a podnebí shrnujeme v přehledu:
- zeměpisná šířka: 49°56'47"
- zeměpisná délka: 14°10'33"
- nadmořská výška: 365 m
- roční průměr srážek: 520 mm
- průměrná teplota roku: 8,3 °C
- průměrná teplota července: 18,0 °C
- průměrná teplota ledna: −1,2 °C
- průměrná teplota vegetačního období: 14,8 °C
Oble vyklenutý hřbet Skalky tvoří antiklinála chotečských vápenců středního devonu, která se noří ze srbských břidlic (Svoboda, Prantl 1955) budujících nižší plochý terén v s., v. a zejména j. sousedství, kde se táhne rozlehlá enkláva polí uvnitř lesů, pomístně označovaná jako Čihová. Jelikož se název Skalka opakuje na různých místech, označujeme zmíněné návrší jako Skalka nad Čihovou. Její svahy jsou ve spodním úseku poněkud prudší, zejména pak na s. úbočí, kde v této úrovni vystupují i drobné skalky se suťovými osypy. V první z nich směrem od sv. konce Skalky leží popisovaná jeskyně (mapka).
Svah nad jeskyní pokrývá dubohabrová pařezina, která na kamenitých místech přechází do útržkovitě vyvinutých porostů habru, mléčného javoru a lípy, kde se v bylinném patře uplatňuje bažanka (Mercurialis perennis L.). Výše ve vrcholové části hřbetu a na j. svahu se objevuje i dřín. Pod jeskyní na mírně ukloněné rovině jsou nové výsadby, zčásti smrkové, které vznikly v nedávné době na bývalých polích, tvořících zde na břidlicích výběžek zasahující k západu do lesního celku.
V současné době jeskyni z velké části zaplňují sedimenty, takže výška stropu nad výplní dosahuje jen 20—40 cm. Výplň se mírně sklání, jak směrem do nitra jeskyně, tak směrem před vchod, takže tvoří plochý val. Jeho vnější část odkryl výkop, jehož čelní stěna leží těsně před okapovou čarou v nejvyšší části valu. Profil v. a s. (= čelní) stěnou výkopu znázorňuje obrázek doplněný podrobným popisem vrstev.
Všechny vrstvy poskytly fosilní ulity, kosti a zuby, poloha 4 i archeologické památky z období eneolitu, které byly předány V. Matouškovi z Okresního muzea v Berouně. Rostlinné fosilie jsou zastoupené roztroušenými uhlíky a některými semeny. Daleko nejbohatší nálezový celek představují měkkýši, jimž v rámci této studie věnujeme hlavní pozornost.
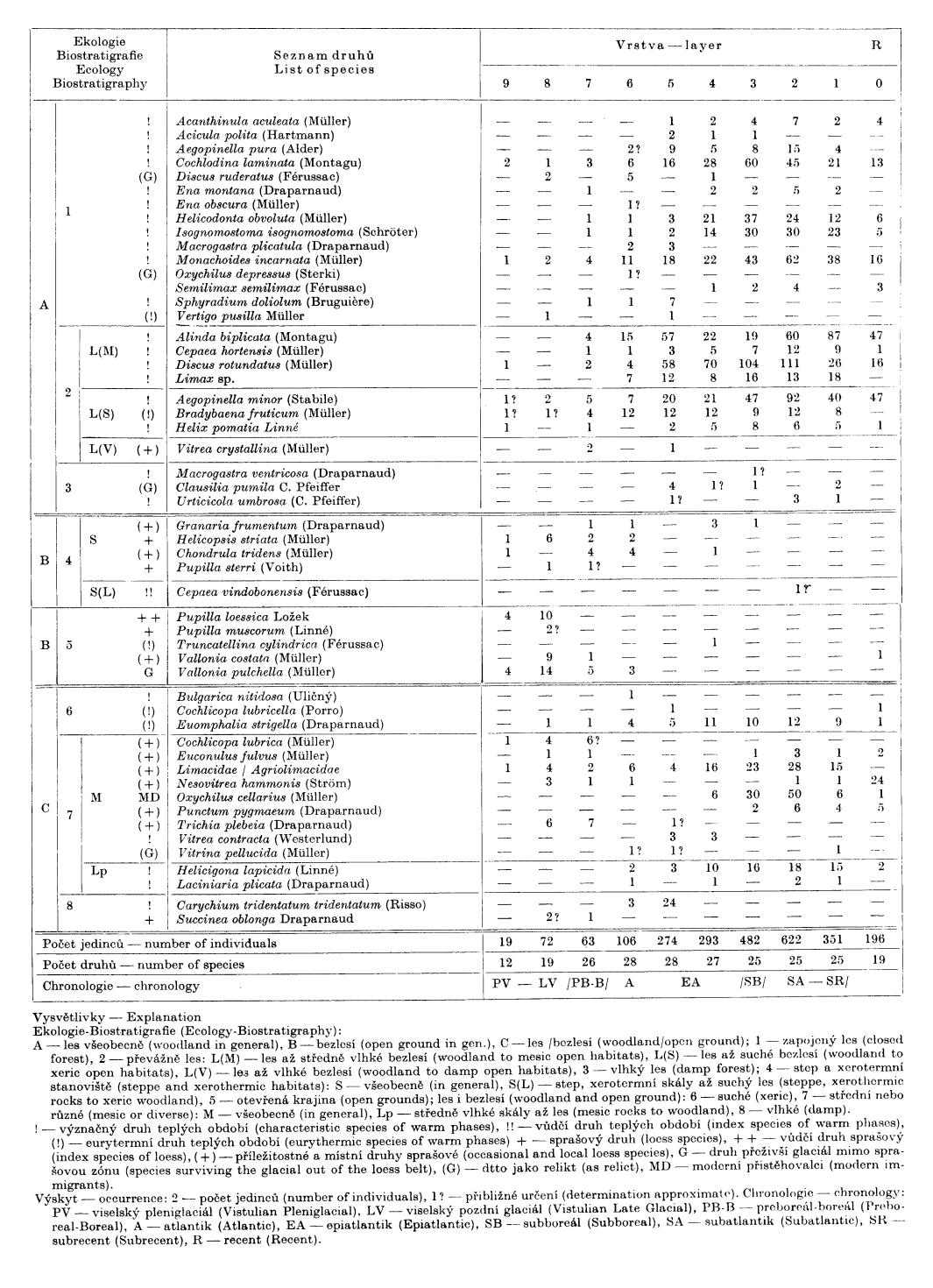
Tabulka 1
Skalka nad Čihovou: měkkýší fauna. – Skalka nad Čihovou: molluscan fauna.
Rozbor malakofauny
Ze všech makroskopicky rozlišitelných vrstev byly odebrány vzorky o přibližném objemu 32 000—35 000 cm3, rozplaveny a vybrány. Přes poměrně velký objem však zejména spodní úsek poskytl jen poměrně chudý materiál, který nedovoluje spolehlivé statistické zpracování ani grafické vyjádření malakodiagramem.
Výsledky sběru zpracované jednotnou metodikou (Ložek 1964) shrnuje tabulka 1 se základními údaji o paleoekologickém a biostratigrafickém významu jednotlivých druhů. Z grafických příloh zařazujeme jen spektra s poukazem, že jejich spodní úsek (zvláště vrstvy 9 a 8) je zřejmě zkreslen vzhledem k malému obsahu ulit. Nicméně i tento materiál umožňuje některé významné závěry, jak vyplývá z následujícího rozboru (srv. Ložek 1982b).
Bazální souvrství hrubých sutí a balvanů, jejichž výplň tvoří ulehlá provápněná jemnozem ± sprašovitého rázu (9, 8), chová jen chudou faunu se zřetelnou převahou druhů otevřené krajiny (B), popřípadě indiferentů (C). Početně nedostatečný soubor nálezů poskytla zejména nejhlubší dosažená poloha 9, kde se výrazněji uplatňují jen Pupilla loessica a Vallonia pulchella. Celkový obraz narušují některé lesní druhy, zastoupené ovšem jen jednotlivými zlomky ulit. Většinou asi jde o úlomky zavlečené z nadloží, jak tomu nasvědčuje i jejich fosilizace. S poněkud výstižnějším obrazem se setkáváme ve vrstvě 8, kde se sice ještě udržuje výrazný podíl pleniglaciálního prvku P. loessica, zároveň však nastupují i indiferenti včetně Trichia plebeia, která je v Českém krasu význačná pro starý holocén. Z celkového charakteru fauny i litologického složení vrstev je zřejmé, že souvrství odpovídá samému konci pleistocénu. Společenstvo vrstvy 8 má pozdně glaciální ráz, z něhož se vymyká jen vysoký podíl P. loessica. Podložní vrstva 9 je natolik chudá, že paleomalakologicky nelze bezpečně prokázat její vznik na konci pleniglaciálu, i když je to nejvýš pravděpodobné.
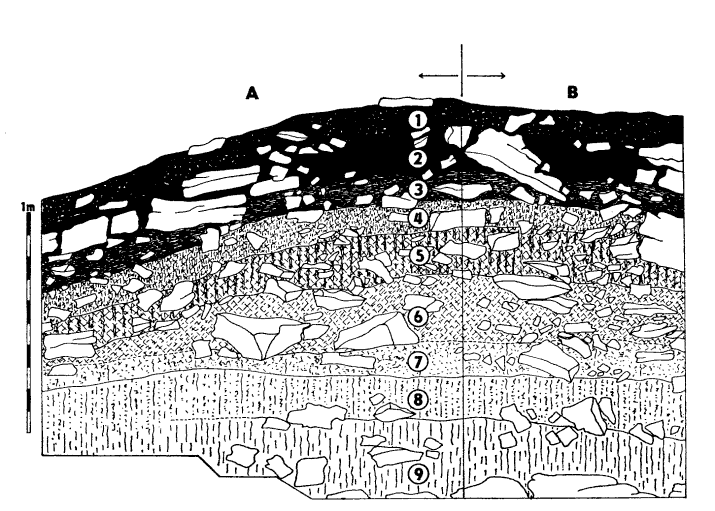
Skalka nad Čihovou. Usazeniny ve vchodu jeskyně: 1 – černá (7,5 YR 1/1) humózní hlína s většími kameny i drobnou drtí; 2 – šedavě černá (7,5 YR 1,5/1) humózní hlína s hrubými bloky, téměř bez drobné drti; 3 – hnědavě tmavošedá (7,5 YR 2,5/1) humózní hlína s hrubými bloky i menšími oleptanými úlomky; 4 – šedavě hnědá (7,5 YR 3/2) jílovitá hlína s většími kameny i menšími úlomky, humózní infiltrace, keramika; 5 – narudle hnědá (7,5 YR 3/4) jílovitá hlína s hrubšími kameny i drobnou drtí; 6 – sytě hnědá (7,5 YR 4/4) jílovitá hlína s hrubými deskovitými bloky i drobnou drtí; 7 – narudle hnědá (7,5 YR 3,5/4) jílovitá hlína se světlejšími provápněnými partiemi s velkými kameny i střední drtí; 8 – žlutohnědá (10 YR 4/7) provápněná hlína s bloky a hojnou drobnou drtí; 9 – světleji žlutohnědá (10 YR 5/6) ulehlá sprašovitá hlína s velkými bloky a drobnou drtí. – A – východní, B – čelní úsek profilu.
Skalka nad Čihovou. Deposits in the cave entrance: 1 – black (7,5 YR 1/1) humic loam with larger fragments and fine debris; 2 – greyish black (7,5 YR 1,5/1) humic loam with coarse fragments, very poor in fine debris; 3 – dark brownish grey (7,5 YR 2,5/1) humic loam with boulders and fine corroded debris; 4 – greyish brown (7,5 YR 3/2) clay loam with coarse fragments and fine debris, humus infiltrations, pottery; 5 – strong brown (7,5 YR 3/4) clay loam with coarse fragments and fine debris; 6 – brown (7,5 YR 4/4) clay loam with coarse flat boulders and fine debris; 7 – reddish brown (7,5 YR 3,5/4) clay loam with highter calcareous parts (becoming increasingly calcareous and paler with depth), boulders, medium sized debris; 8 – dark yellowish brown (10 YR 4/7) calcareous loam with boulders, rich in fine debris; 9 – yellowish brown (10 YR 5/6) loess-like loam, condensed and forming matrix of very coarse breakdown, rich in fine debris. – A – eastern, B – front sector of the section.
Vrstva 7 se rovněž vyznačuje malým počtem nálezů, nicméně druhové bohatství vzrůstá. I zde je silně zastoupena stepní složka, kde k druhům Helicopsis striata a Chondrula tridens přistupuje i Granaria frumentum. Trichia plebeia podržuje svůj podíl, význam má dále nástup druhu Vitrea crystallina a již výrazné zastoupení Bradybaena fruticum. Podstatně vyšší je i podíl lesních prvků (A), mezi nimiž se ve vyšších počtech objevují časně holocenní přistěhovalci, jako Aegopinella minor nebo Cochlodina laminata. Řada zlomků je silně korodována, takže jsou většinou neurčitelné, jiné jsou však dobře zachovalé. Tomu odpovídá i celkový vývoj vrstvy, kde se střídají partie provápněné s úseky v různé míře odvápněnými. Jde tedy o polohu přechodnou, se smíšenou faunou, v níž se projevuje postupný nástup lesních prvků. Tento obraz odpovídá starému holocénu, tj. preboreálu a boreálu (malakozóna C – Ložek 1982b).
Vrstva 6 je již o něco bohatší, stále však ji lze ještě označit jako poměrně chudou, i když zde hraje svou úlohu vysoký podíl neurčitelných korodovaných zlomků, které nejsou započítány. Druhová diverzita zde vrcholí, což podmiňuje jednak doznívající výskyt stepních druhů, jednak nástup dalších náročných, především lesních prvků. Význačné je maximum druhu Discus ruderatus, nástup Macrogastra plicatula a Carychium tridentatum, které se dnes váží na údolní zářezy, i dalších druhů, jako Helicigona lapicida a Laciniaria plicata. Výhradně zde se objevuje Bulgarica nitidosa, která dnes v těchto polohách rovněž nežije, ač je jinak v Českém krasu poměrně hojná. Dále zaslouží zmínky silný vzestup Alinda biplicata i významné zastoupení druhů Bradybaena fruticum a Euomphalia strigella, které ukazuje, že porosty byly dosud světlé a že se uplatňovaly plášťové formace. V této vrstvě s konečnou platností nabývá převahy zapojený les a výrazně se projevuje zvlhčení. Druhy otevřené krajiny postupně mizí. Tento obraz, význačný vysokým stupněm stanovištní diverzity, odpovídá atlantiku v pojetí K.-D. Jägera (1969).
Ve vrstvě 5 se zachovává vysoké druhové bohatství. Místo vymizelých obyvatel otevřených ploch se objevuje řada nových lesních prvků, jako Acanthinula aculeata, Acicula polita, včetně vlhkomilné Clausilia pumila a patrně i Urticicola umbrosa. Macrogastra plicatula se udržuje, Carychium tridentatum dosahuje maxima. Rozvoj lesního ekosystému zde vrcholí, což odpovídá epiatlantiku (malakozóna E – Ložek 1982b).
Vrstva 4 sice vykazuje obdobné poměry, nicméně mizí druhy citlivé na vysušení, jako Carychium tridentatum a Macrogastra plicatula, a znovu se objevují stepní prvky Granaria frumentum a Chondrula tridens včetně jediného výskytu xeroterma Truncatellina cylindrica v celém profilu i vysokého podílu Euomphalia strigella. Vzdor tomu nastupuje Semilimax semilimax. Složení fauny ukazuje na prosvětlení a vysušení, které mohou být nejspíše podmíněné osídlením doloženým v této vrstvě. Vrstvou 4 končí období maximálního druhového bohatství i vlhkosti. Složení malakofauny odpovídá ještě epiatlantiku, což se shoduje s výpovědí archeologických nálezů.
Nadložní souvrství humózních sutí 3—1 se vyznačuje výrazným vzestupem počtu jedinců. Souvisí to s vytvořením volných sutí i s okolností, že většina zlomků je určitelná – tedy s nižším stupněm koroze. Fauna nabývá rychle zhruba současného složení, i když zůstává druhově bohatší. Pro vrstvu 3 je význačná kulminace druhů Helicodonta obvoluta, Cochlodina laminata a Aegopinella pura, zatím co v poloze 2 dosahují vrcholu Discus rotundatus, Aegopinella minor a Cepaea hortensis. Alinda biplicata zato vrcholí až v povrchové poloze 1 a své vysoké zastoupení spolu s druhem Aegopinella minor podržuje dodnes. Složení společenstev je typicky mladoholocenní (malakozóna F). Bližší členění lze provést jen přibližně tím, že vrstvu 3, kde se v plné míře rozvíjí suťové společenstvo, můžeme v souladu s pozorováními na četných jiných lokalitách zařadit do subboreálu a nadložní vrstvy 2 a 1 do staršího a mladšího subatlantiku (= subrecentu). Pokles druhového bohatství, který se projevuje ve složení dnešní fauny, podmínily zřejmě lidské zásahy v nejmladší minulosti, např. existence obdělaných polí v bezprostředním sousedství jeskyně. Dnes se spíše projevuje opětná regenerace lesních společenstev.
Z malakozoologických nálezů zasluhují zvláštní zmínky:
- Vallonia pulchella, která se vyskytuje v poměrně výrazném podílu v pozdně pleistocenním úseku, představuje v tomto vyloženě suchém prostředí poměrně vzácný nález, neboť se v glaciálech většinou váže na mokřadní stanoviště, což však neplatí pro dnešek.
- Laciniaria plicata se v pozdější fázi klimatického optima ojediněle vyskytuje na větším počtu středočeských nalezišť, kde dnes již nežije. Zde však přežívá i do nejmladšího úseku, což je rovněž výjimečné.
- Bulgarica nitidosa se v klimatickém optimu objevuje i tam, kde se dnes již nevyskytuje. Ojedinělý nález ve vrstvě 6 je dalším potvrzením tohoto pravidla.
Jinak vývoj malakofauny zachycený v profilu na Skalce představuje celkem typický sled postglaciálních společenstev, který je zde vzhledem k celkovým stanovištním poměrům (nedostatek mokřadních i skalních prvků) poměrně stejnorodý, což má výhodu v tom, že zde určité, dočasně přemnožené druhy nenarušují relativní početní zastoupení.
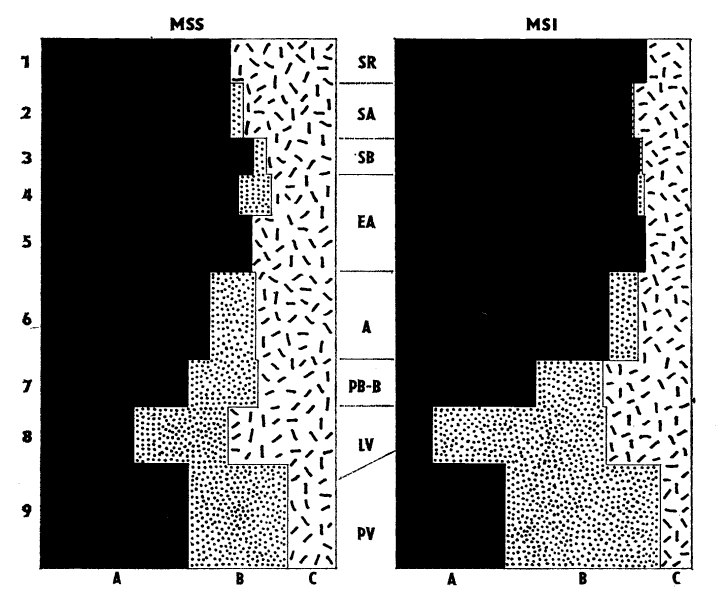
Malakospektra druhů (MSS) a jedinců (MSI) vyjadřující relativní zastoupení hlavních ekologických skupin (A—C, viz tab. 1). Vlevo čísla vrstev, uprostřed chronologie (viz tab. 1).
Malacospectra of species (MSS) and inidividuals (MSI) showing the percentage of main ecologic groups (A—C, sp. tab. 1); numbers of layers on the left, chronology in the middle column (cp. tab. 1).
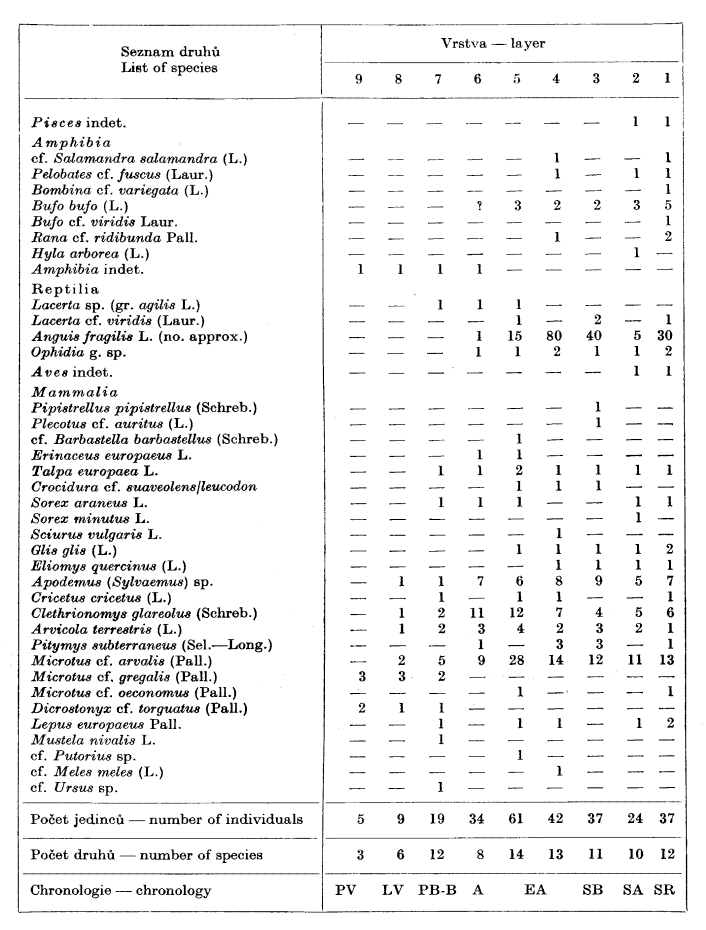
Obratlovci (Ivan Horáček)
Pozůstatky obratlovců vybrané ze vzorků společně s měkkýši sice nevynikají četností, zachycují však přesto poměrně velký počet druhů, jak ukazuje přehledná tabulka 2. Pozoruhodný je bohatý výskyt různých druhů obojživelníků včetně rosničky a blatnice.
Výskyt klimaticky a stanovištně významných druhů je v dobrém souladu s již zmíněnými výsledky malakozoologického rozboru, což názorně ukazuje souborné vyhodnocení celého profilu na tabulce 3.
Tabulka 2
Skalka nad Čihovou: obratlovci. – Skalka nad Čihovou: vertebrates.
Stratigrafické a paleogeografické poznatky
Stanovištní vývoj na Skalce lze odvodit z podrobného rozboru měkkýší fauny, která dostatečně charakterizuje jeho jednotlivé fáze. Korelaci malakologických poznatků s ostatními pozorováními shrnuje souborné vyhodnocení (tab.3), z něhož vycházejí širší úvahy probírané v této kapitole.
Profil na Skalce odkryl typickou ukázku trojdílného postglaciálního souvrství, v němž lze sledovat nejen změny sedimentace, ale i intenzitu půdotvorných pochodů (Kukla, Ložek 1971). Z biostratigrafického rozboru je zřejmé, že pedogeneze byla nejsilnější v době klimatického optima, což souviselo s vyšší vlhkostí, která se zřetelně obráží i ve složení malakofauny. Jemnozem příslušných vrstev, (5, 6 a zčásti i 7) je v různé míře odvápněna a prosycena uvolněným jílem. Odvápnění se projevuje korozí mnoha ulit, takže poměrně vysoký podíl plžů je neurčitelný. To svědčí pro poměrně klidnou sedimentaci, kdy se uvolňovalo jen menší množství drobných vápencových úlomků a tvořící se sediment průběžně postihovalo zvětrávání vlivem půdotvorného pochodu, jehož produktem je silně vyvinutá vápencová hnědozem až slabá terra fusca. Pod skalním srubem ve vchodu jeskyně byl tento pochod ovšem neustále rušen opadem vápencového materiálu z portálu, takže zde nedosáhl plné intenzity.
Profily obdobného vývoje jsou známé z četných míst v Českém krasu. Na prvním místě lze uvést odkryv v lůmku proti skále Zubáku v rokli pod Zadní Kopaninou, kde klimatickému optimu odpovídá zcela odvápněná, a tedy paleozoologicky sterilní vrstva, ačkoli podloží i nadloží jsou vápnité a obsahují fosilní ulity. Jde o úpatní osyp, kde se půdotvorní činitelé mohly silněji uplatnit než v portálu jeskyně. Profil představuje typickou ukázku tohoto vývoje a byl jako takový uveřejněn (Ložek 1973) a předveden holocenní komisi INQUA (Ložek in Kliewe 1967). Další obdobou je dosud neuveřejněný profil ve vchodu jeskyně Martina u Tetína, kde nejsilněji odvápněná poloha leží v bezprostředním podloží vrstvy s neolitickou kulturou lineární (volutovou). Podobný vývoj mají i sedimenty před portálem Rudolfovy jeskyně v Přední rokli u Karlštejna. Méně výrazná je příslušná poloha v převisu na západním svahu Kobyly u Koněprus, zatímco tam, kde v okolí trvale převládaly humózní rendziny, může být celé holocenní souvrství tvořeno jen silně vápnitými rendzinovými sedimenty (Axamitova brána na Kotýzu).
Stratigraficky odpovídají hnědé, více méně odvápněné horizonty polohám pěnitce, který se ovšem může tvořit jen pod skalní klenbou ze skapávající vody (Ložek 1984, 1985). Platí to i pro starší teplá období, jak nejlépe dokládá vrstevní defilé odkryté na boku Stránské skály, kde parahnědozem sprašové série přechází pod skalní stěnou do sypkých sintrů – pěnitců (Ložek 1965). Přitom jde o interglaciál, jehož staropleistocenní stáří dokládá biharská fauna.
Shrneme-li zmíněné poznatky, je zřejmé, že i v souvrstvích z čistě klastických materiálů lze sledovat dva hlavní výkyvy vlhkosti během holocénu: 1. starší, mimořádně vlhké fázi, doložené tvorbou pěnitce ve vchodech jeskyní a pod převisy, odpovídají hnědozemní sedimenty s částečně až úplně odvápněnou jemnozemí, zatímco 2. mladší, výrazně suchý výkyv, jehož hlavním dokladem jsou pohřbené rendziny v pěnovcích, vedl k rychlé tvorbě hrubší suti, často jen nedokonale vyplněné vápnitou humózní jemnozemí (v typickém vývoji např. v Přídolí u Velké Chuchle nebo v profilu Srbsko – Hřiště).
Skalka nad Čihovou podává svědectví i o průběhu odnosných a sedimentačních pochodů v nejmladší geologické minulosti. I v této poloze – v úrovni jen mírně snížené paroviny, kde je možnost výskytu i velmi starých pokryvů (křída, terciér), tvoří osyp z valné části sedimenty, které se usadily v období od posledního (viselského) pleniglaciálu do současné doby. Stopy starších uloženin zde zjištěny nebyly, takže i zde lze počítat s působením silného odnosu ve starší fázi posledního pleniglaciálu, které odpovídá ústup svahů včetně jeskynních vchodů (Ložek 1980) a přibližně i poslední fáze silné hloubkové eroze (Ložek 1981). Vzhledem k málo svažitému terénu nutno vzít v úvahu především periglaciální pochody, zejména soliflukci.
Horizonty jednotlivých holocenních fází vykazují na Skalce poměrně rovnoměrný vývoj co do mocnosti, čímž se liší od většiny postglaciálních souvrství v údolních zářezech nebo v exponovaných polohách strmějších svahů, kde se uplatnila dílčí erozní fáze, která vedla k tomu, že hlavní mocnost sedimentace buď spadá do staršího holocénu a pozdního glaciálu, nebo naopak až do středního a mladého holocénu. Toto rozlišení postglaciálních souvrství do dvou skupin různého vývoje svědčí o tom, že uvedená eroze dočasně působila někdy na rozhraní staršího a středního holocénu, nejspíše na počátku atlantiku (Jäger, Ložek 1983). Vzhledem k tomu, že rovnoměrný vývoj celého postglaciálního souvrství, ovšem se silnými stopami pedogeneze ve střední části, nacházíme především v plošších polohách mimo čerstvé údolní zářezy (Martina, Kobyla ap.), lze soudit, že tato mladší – holocenní eroze zasáhla především jen údolní polohy a nebyla znásobena intenzívním plošným odnosem, podmíněným periglaciálními poměry, jako tomu bylo v případě zmíněné starší odnosné fáze ve starším pleniglaciálu.
Hrubší sutě pod svahy Skalky i dalších podobných vyvýšenin v sousedství jsou tedy zřejmě velmi mladého, tj. mladoholocenního původu, stejně jako je tomu u volných osypů na úpatí údolních svahů [Srbsko – Hřiště (Ložek 1969), Srbsko – Kužel (Ložek 1982a), Přidolí (Ložek, Kukla 1956) aj.]. Je to dokladem, že mladoholocenní oživení fyzikálního zvětrávání a přesunu jeho produktů zasáhlo i parovinné polohy, a že tedy nejde o zvětrávání periglaciální, jak se někdy předpokládá (Ložek 1963).
Tabulka 3

Význam zjištěných poznatků pro ochranu přírody
Skalka nad Čihovou leží ve státní přírodní rezervaci „Karlštejn“, která patří mezi naše nejvýznamnější chráněná území. Zahrnuje ovšem jak plochy dosud blízké přírodnímu stavu, tak okrsky změněné hospodářskými zásahy. K nim patří i Skalka, kterou dnes kryje nízký výmladkový les jako většinu okolních pahorků podobného rázu. Ze srovnání současné fauny se společenstvy, která zde žila v minulosti, vidíme, že se dnešní stav vyznačuje zřetelným ochuzením. To nepochybně souvisí s bezprostředním sousedstvím polí, která donedávna zasahovala až těsně pod jeskyni, i s obhospodařováním lesního porostu. Cílem ochrany je ovšem navodit stav, který by se více přiblížil někdejším přírodním poměrům a podmínil plný rozvoj místních biocenóz. Tyto záměry je třeba opřít o znalost stanovištního vývoje v nejmladší minulosti, zejména ve vztahu k případným lidským zásahům.
Z biostratigrafického rozboru je zřejmé, že vývoj ekosystémů na s. svahu Skalky probíhal poměrně klidně bez náhlých zvratů. Dokládá to plynulý vývoj malakofauny od chudých pozdně glaciálních společenstev otevřené krajiny do bohatě rozvinutých malakocenóz klimatického optima (vrstvy 6 a 5), která zahrnují i druhy dnes v tomto prostoru vymřelé. Od tohoto vrcholu se směrem k dnešku projevuje mírné ochuzení, vrcholící v poměrně nedávné době. Náznaky určitého narušení se projevují jen ve vrstvě 4, obsahující eneolitické památky, a to dočasným výskytem několika stepních druhů (Granaria, Chondrula, Truncatellina). Lze předpokládat, že za tohoto osídlení rozvolnila les na Skalce pastva, takže zde vznikly stepní světliny, jaké se vyskytují dodnes na některých okolních kopcích, např. na Mokrém vrchu nebo Koníčku. Ty však později opět zarostly, což podmínilo opětné vymizení stepních druhů.
Cílem ochrany by měl být přibližně takový stav fauny i celého stanoviště, jako se jeví ve vrstvě 2 a do určité míry i v povrchové poloze 1. Je nepochybné, že vytvoření pařezin vedlo k ochuzení malakofauny, zejména o druhy, které nejlépe prospívají na padlém dřevu. Druhy žijící v suti jsou postiženy jen při hrubých zásazích, jako je delší odlesnění a vysušení, k čemuž zde však zřejmě nikdy nedošlo. Výběžek poli, který se táhl přímo pod jeskyni a podle různých náznaků pokračoval v hlubší minulosti nesouvisle směrem ke Králově studni, je dnes zalesněn, čímž se zlepšilo porostní klima při jeskyni a lesní fauna má lepší možnost rozvoje než ještě před nedávnem. Nahrazení pařeziny vysokým lesem, aspoň na strmém severním svahu, by nepochybně umožnilo další rozmach lesního společenstva a snad i návrat některých druhů, které zde dnes již nežijí. Je proto třeba zvážit postupný převod aspoň části pařezin na kmenoviny – ovšem bez větších změn ve složení porostů. Tím by se nastolil opět stav blízký přírodním poměrům, což je v tomto prostoru možné vzhledem k tomu, že během minulosti zde nedošlo k tak negativním zásahům, aby změny ekosystémů byly nevratné.
* * *
Z biostratigrafického rozboru vstupního valu jeskyně na Skalce nad Čihovou lze odvodit tyto poznatky:
- Souvrství chová plynulý sled měkkýších společenstev od konce viselského pleniglaciálu do současné doby.
- Ve sledu malakocenóz lze rozlišit 3 fáze: období chudých společenstev z konce glaciálu, v nichž se během staršího holocénu postupně prosazují lesní druhy; maximum lesních a na vlhko náročných druhů v době klimatického optima a mírně ochuzené, dnešním společenstvům obdobné fauny v mladém holocénu.
- Tři zřetelné fáze jeví i sedimentace: sprašovité a silně vápnité, humusem chudé hlíny na bázi, hnědé hlíny s uvolněným jílem a zčásti odvápněnou jemnozemí ve středu a černé kypré vápnité hlíny se sutí v horní části profilu.
- Starší sedimenty zde nebyly zjištěny, neboť zřejmě byly vyklizeny v době poslední velké odnosné fáze ve starším viselském pleniglaciálu.
- Celý profil je rovnoměrně vyvinut, což nasvědčuje, že nebyl silněji postižen dílčí erozí během holocénu.
- Hlavní vlhký výkyv holocénu se projevuje dílčím odvápněním jemnozemi středních členů souvrství v atlantiku, suchý výkyv subboreálu vedl k tvorbě sutí s kyprou vápnitou humózní výplní.
- Pedogenetické pochody spjaté s intenzívním přesunem CaCO3 v roztoku se nejsilněji projevují v době klimatického optima.
Literatura:
- Jäger K.D. (1969): Climatic Character and Oscillations of the Subboreal Period in the Dry Regions of the Central European Highlands. – Proceedings of the VII Congress INQUA, 16: 38—42. Washington.
- Jäger K.D., Ložek V. (1983): Paleohydrological implications on the Holocene development of climate in Central Europe based on depositional sequences of calcareous fresh-water sediments. – Quaternary Studies in Poland, 4: 81—89. Warszawa, Poznań.
- Kliewe H. et al. (1967): Probleme und Berunde der Holozänstratigraphie in Thüringen, Sachsen und Böhmen. – Quartärkomitee der DDR: 1—180. Berlin, Prag.
- Kukla J., Ložek V. (1971): Význam krasových oblastí pro poznání poledové doby. – Československý kras, 20 (1968): 35—49. Praha.
- Ložek V. (1963): K otázce tvorby svahových sutí v Českém krasu. – Československý kras, 14 (1962—1963): 7—16. Praha.
- Ložek V. (1964): Quartärmollusken der Tschechoslowakei. – Rozpravy Ústředního ústavu geologického, 31: 31—374, 36 příloh. Praha.
- Ložek V. (1965): The relationship between the development of soils and faunas in the warm Quaternary phases. – Sborník geologických věd, řada Antropozoikum, 3: 7—33, 3 přílohy. Praha.
- Ložek V. (1969): Značenije molljuskov dlja izučenija kontinentaľnogo golocena. – Golocen (k VIII Kongressu INQUA), Paris 1969: 58—78. Moskva.
- Ložek V. (1973): Příroda ve čtvrtohorách. – Academia: 1—372, 26 příloh. Praha.
- Ložek V. (1974): Příroda Českého krasu v nejmladší geologické minulosti. – Bohemia Centralis, 3: 175—194, 2 přílohy. Praha.
- Ložek V. (1980): Chronological position of the last phase of slope retreat in Czechoslovak karst areas. – Československý kras, 31 (1979): 7—17. Praha.
- Ložek V. (1981): Zur Altersstellung der jüngsten Tiefenerosion in den Tälern der innerböhmischen Hügelländer. – Biuletyn Instytutu Geologicznego, 321: 239—248. Warszawa.
- Ložek V. (1982a): Faunengeschichtliche Grundlinien zur spät- und nacheiszeitlichen Entwicklung der Molluskenbestände in Mitteleuropa. – Rozpravy Československé akademie věd, řada matematických a přírodních věd, 92/4: 1—106, 11 příloh. Praha.
- Ložek V. (1982b): Contribution of Malacology to the Chronological Subdivision of the Central European Holocene. – Striae, 16: 84—87. Uppsala.
- Ložek V. (1984): The Foam Sinter as Palaeoclimatic Indicator. – Československý kras, 34: 7—14. Praha.
- Ložek V. (1985): The site of Soutěska and its significance for Holocene climatic development. – Československý kras, 36: 7—22, 1 příloha. Praha.
- Ložek V., Kukla J. (1956): Profil holocenními svahovinami ve Velké Chuchli (V Dolích). – Anthropozoikum, 5: 407—423, 1 příloha. Praha.
- Svoboda J., Prantl F. (1955): O stratigrafii a tektonice staršího paleozoika v širším okolí Karlštejna. – Sborník Ústředního ústavu geologického, Oddíl geologický, 21/1 (1954): 519—596, 5 příloh. Praha.
Summary:
The cave site of Skalka nad Čihovou near Karlštejn in the Bohemian Karst was investigated in 1984. In contrast to the great majority of cave sites in this area which lie in deep valley cuts or in steep rocky slopes the Skalka Cave is situated at the foot of the north-facing slope of a low hill consisting of Devonian limestones and forming a part of the undulated mid-Bohemian peneplain. In comparison with valley cuts and rocky slopes the environmental conditions at Skalka are characterized by a low diversity and largely correspond to those of climax ecosystems within the area under study. Thus a biostratigraphic analysis of the cave fill can provide data on their faunal development in relation to accumulation, pedogenesis and erosion which have poorly been known until now. The results may be utilized in managing the platform sectors of the Nature Reserve „Karlštejn“ to which the Skalka area belongs.
Basic data on the topography and climate of the site investigated are listed below:
- Latitude: 49°56'47"
- Longitude: 14°10'33"
- Altitude: 365 m
- Mean annual rainfall: 520 mm
- Mean annual temperature: 8,3 °C
- Mean temperature of January: −1,2 °C
- Mean temperature of July: 18,0 °C
- Mean temperature of vegetational period: 14,8 °C
The slope of Skalka above the cave is covered by a low Querco-Carpinetum, whereas on small rocky outcrops and scree formations at the cave entrance level patches of a Tilio-Acerion occur. On the top of Skalka and on its south-facing slope the tree canopy is more xeric and includes e.g. Cornus mas L. and other thermophilous elements.
The section was exposed by a pit dug in sediments accumulated in the cave entrance. Its development with the description of particular layers are given in the figure on pg. 59. Samples of ca 32 000—35 000 cm3 were taken from all layers, air dried, wet sieved and the shells and bones extracted. The standard techniques of sampling, extraction and shell counting are described by V. Ložek (1964). The results are given in table 1 (Mollusca) which includes also the basic information on the ecology and biostratigraphy of each species. The Vertebrate remains are listed in table 2.
The deposits consist of coarse breakdown coming from the gradually destroyed cave entrance and of loamy matrix which enables the column of the excavation to be subdivided into three main units containing also characteristic faunal assemblages. A review of all results is given in table 3 (pg.65).
The basal layers 9 and 8 have a loessoid matrix and include a poor tolerant open ground fauna with Pupilla loessica, Dicrostonyx and Microtus gregalis. For this reason they are attributable to the latest Pleistocene, i.e. to the decline of the Vistulian Pleniglacial (9) and to the Late Vistulian (8). The occurrence of sparse fragments of less tolerant snail species may be due to a later contamination from the overlying strata.
Also the layer 7 is dominated by an open ground fauna in which the xerothermic Granaria frumentum appears. Nevertheless, the woodland elements occur in higher numbers and many shells are strongly corroded which corresponds to a partial decalcification of the fine earth, i.e. to an increase of pedogenesis. Hence, this layer has a transitional position and can be assigned to the Early Holocene (Preboreal-Boreal).
The layer 6 is characterized by the last occurrences of xerothermic step pe elements (Chondru la, Granaria) and by an invasion of demanding woodland species as Macrogastra plicatula. Alinda biplicata, Apodemus (Sylvaemus) and Clethrionomys become important, but Discus ruderatus is still present. The zoocoenosis is very rich in species and shows a very high ecological diversity which is characteristic of the Atlantic sensu JÄGER (1969).
In the overlying layers 5 and 4 the situation is similar: in 5 the woodland fauna attains its optimum, whereas in 4 several open ground elements (Granaria, Chondrula, Truncatellina) temporary appear which may be linked with a forest clearance due to human impact (presence of Eneolithic pottery). The absolute dominance of woodland fauna and the culmination of Carychium tridentatum in the layer 5 correspond to the Epiatlantic (= malacozone E – Ložek 1982b).
The shell corrosion due to partial decalcification of the fine earth can be traced throughout the complex 6—4 and corresponds to intense pedogenetic processes which are only partly hindered by fresh calcareous breakup coming from the surrounding bedrock. The layer 4 can be regarded as transitional.
The upper unit 3—1 differs from the underlying ones in its matrix which is very loose, humic and more calcareous. This is particularly true of the layers 3 and 2 which were probably deposited as a scree poor in matrix providing favourable conditions for such species as Helicodonta obvoluta, Isognomostoma isognomostoma, Discus rotundatus or Alinda biplicata which are found in high numbers in present-day scree formations within the adjacent area. This development is characteristic of the Late Holocene. The depauperization of the recent molluscan fauna is due to the human impact (existence of fields in the nearest vicinity).
The sequence under study developed under purely terrestrial conditions, which must be taken into consideration in its palaeoclimatic interpretation (particulalry focused upon moisture fluctuations). The intense increase of moisture towards the end of the Boreal and in the early Atlantic documented by the precipitation of foam sinter in caves or rock-shelters (Ložek 1984, 1985) corresponds in the Skalka Site to an intense decalcification starting in the layer 7 and culminating during the formation of 6. Analogous processes can be observed in a number of sites in the Bohemian Karst (Martina, Přední rokle, Kobyla). In the foot-slope section near Zadní Kopanina the horizon in question is formed by a completely decalcified brown soil (Ložek in Kliewe 1967).
The temporary decrease of moisture corresponding to the Late Bronze Age (Subboreal sensu Jäger 1969) which induced the break in tufa sedimentation manifested in the occurrence of burried rendzina horizons, can be traced in Skalka and similar sites as a facies change: scree with condensed and partly decalcified matrix is replaced by rapidly accumulated scree with loose humic and more calcareous matrix (Skalka) or in certain cases with free interstices (e.g. Přídolí – Kukla, Ložek 1956, Srbsko-Hřiště – Ložek 1969).
Although the site of Skalka is situated on a platform where very old deposits may occur, the fill of the cave entrance include deposits of Late Pleistocene and Holocene age only. Thus we can suppose that also in these contexts the last phase of intense erosion during the early Vistulian Pleniglacial took place having removed older deposits (Ložek 1980). In comparison with depositional sequences situated in valleys or in steep slopes in which either the Late Glacial-Early Holocene or the Middle-Late Holocene sectors commonly are reduced due to an erosional phase at the Early/Middle Holocene boundary (Jäger, Ložek 1983), all horizons in Skalka are rather equally developed. This is true also of other sites in similar geomorphic contexts and supports the opinion that this erosional phase did not affected the deposits in platform environments.
The aims of our study were to examine the Postglacial pattern of cave entrance sedimentation and faunal development in peneplain environments with low habitat diversity which took place under purely terrestrial conditions largely corresponding to the local climax.
